

|
|

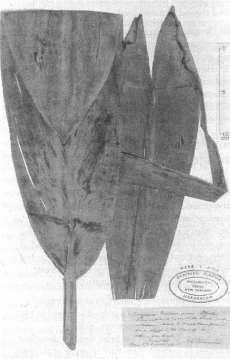 |
| Fig. 1 Herbarium specimen of what is considered to be the last survivor of Cordyline fruticosa (ti pore) introduced by Mäori. It was collected by Thomas Kirk at Ahipara in 1893 and thought to be a new species (Draceana Cheesemanii) before being recognised as C. terminalis, a synonym for C. fruticosa. |
Original horticultural use of Cordyline
Probably the first domestication of Cordyline was in New Guinea. This was for Cordyline fruticosa (syn. C. terminalis). Coloured-leaf variants of this species still abound in Papua New Guinea. The Polynesian in their migrations dispersed this plant throughout the Pacific. It was one of the six plants that Maori introduced to Aotearoa known to have survived in cultivation at least until the time Cook visited in 1769.
The last of the C. fruticosa introduced by Maori in mainland New Zealand was found at Ahipara (Fig. 1). This plant attracted the excited attention of botanists at the end of last century. Pieces of it were taken and grown in the gardens of these botanists but both these and the Ahipara plant died. In 1900 Canon Philip Walsh recorded most of what we now know about the Maori use of this plant and the name ti pore that they gave to it (Walsh 1900).
Relics of the Polynesian introduction of C. fruticosa can be still found on Raoul Island in the Kermadecs. Bill Sykes brought living specimens of this plant back to New Zealand some years ago and this is still growing at Landcare Research, Lincoln.
Both Cook Islanders and Maori know Raoul Island as Rangitahua. This name translates as 'fire in the sky' indicating that both these people saw its massive volcanic eruptions. Rangitahua is linked to the traditions of both the Kurahaupo and Aotea ocean voyaging canoes that bought the ancestors of Maori to Aotearoa.
It is likely that the Raoul Island plants are of the same stock as the last of the Maori ti pore. To mark this connection, a plant propagated from Rangitahua stock has been planted at the Ahipara Primary School. Although not vigorous, it has survived outdoors at the school for several years.
Maori use and domestication of ti
The Cordyline fruticosa introduced by Maori was marginally adapted to the climate of New Zealand. It could only survive with cultivation in the warmest parts of the country. Maori recognised the affinity of this plant to the native New Zealand species of Cordyline. They gave the generic name ti to the native species, and as far as we know used them for fibre, food, medicinal and ceremonial purposes in much the same way as they used C. fruticosa.
After harakeke-flax (Phormium tenax), the cabbage tree, te kouka, was the most important provider of fibre in Maori traditional life. This fibre, in abundant supply from wild plants, was especially useful in that it was more durable than harakeke when used for fishing lines and nets. Beyond the harvesting of wild plants, several levels of domestication were applied to the New Zealand ti to advance their use as a source of food (Harris and Mann 1994).
In the eastern South Island in particular, te kouka was coppiced on a 3 to 4 year rotation and the harvested stems steamed for 24 hours in large umu to produce the sweetener kauru. The steaming converted the carbohydrate fructan in the stems to very sweet fructose. The material processed in this way was stored dry until the time came to add it to fern root and other foods to improve their palatability. Waikato Maori cultivated ti rauriki, the small grass-like northern cabbage tree (C. pumilio), for the sweet carbohydrate of its rhizomes.
Ti Tawhiti
 |
 |
| Fig. 2 Cordyline 'Ti Tawhiti' growing at the base of a cabbage tree (C. australis). |
Fig. 3 Cordyline 'Ti Tawhiti' (left) and C. 'Karo Kiri'. |
Cordyline 'Ti Tawhiti' (Fig. 2, 3) is a remarkable plant in New Zealand's horticultural history. Evidence points to it being a dwarf non-flowering C. australis that was selected and cultivated by Maori. It was saved from extinction because its dwarf form found favour with European gardeners. This is recorded in the first systematic recording of New Zealand trees and shrubs suitable for cultivation that was made by Potts and Gray (1870). Their observations were based on plants in a garden at Governor's Bay near Lyttleton and they wrote "The Titawhiti, of the Whanganui tribes, grows here very well; it has a dark green leaf, and throws off young plants more freely than some of the other species".
Cordyline 'Ti Tawhiti' was the subject of a learned discussion amongst the leading botanists of New Zealand at a meeting of the Royal Society in Wellington in 1895 (Rutland 1896). Adding to its mystery is the fact that it has never been observed to flower. Thus its exact taxonomic affinity remains uncertain. In garden use it came to be known as Cordyline 'Kirkii' recording the interest the notable 19th Century New Zealand botanist Thomas Kirk had in the plant.
Its origin as a Maori selection was forgotten for most of the 20th century until Peter Heenan and I rediscovered its origins in 1991 (Harris & Heenan 1991). The name 'Tawhiti' is equivalent to 'Hawaiki' and indicates the belief that the plant was introduced to Aotearoa in the ancestral ocean voyaging canoes of Maori. However, it is more probable that the name arose from it being moved around its native land as a domesticated plant.
Peter Heenan went to pains to point out that as a Latinised name 'Kirkii' was inappropriate as a cultivar name, giving the plant the name Cordyline 'Thomas Kirk' (Heenan 1991a). This was before we named the plant Cordyline 'Ti Tawhiti' to recognise its correct and fascinating origin. On a recent visit to a retail nursery in Christchurch I was disappointed to see the plant being sold still with the label C. 'Kirkii'!
Ornamental use of New Zealand cabbage trees
The interest in New Zealand cabbage trees as ornamental plants is well substantiated by Peter Heenan's cultivar checklist (Heenan 1991b). Most of the cultivar differentiation has been within C. australis and this has mostly been for leaf colour variation from the normal green, and for variegation. Variations in habit and of leaf arrangement have been of lesser importance, but it is these characters that differentiate C. 'Ti Tawhiti' from its likely parent species C. australis.
A striking addition to the variation of the form of cabbage tree is provided by a plant discovered in 1983 by Mr Trevor Bayliss, nurseryman, Belfast, Christchurch. This plant has been formally described and named C. australis 'Karo Kiri' (Heenan et al. 1994), was granted New Zealand plant variety rights in 1993, and is available through retail nurseries (Fig. 3). It is characterised by very short rigid leaves with distinct yellow midribs on their lower sides. The cultivar has not flowered.
Cordyline 'Karo Kiri' has very useful phenotypic plasticity. It can be adapted to indoor or outdoor patio pot or garden use. Normally the growth is monopodial and the trunk will continue to elongate to a point where it no longer holds itself erect. With the dead leaves that accumulate along the narrow trunk the plant has a bizarre appearance. Of the plants I have seen growing in gardens most have been like this, and especially when the dead leaves are not removed, the best ornamental qualities of the cultivar are not shown.
Cutting the stem off will induce the formation of new shoots just below the cut. In this way the plant can be sculpted to suit its area of use and to provide variety of form. Grown in full light the trunks will be sturdier, the leaves will be shorter and more rigidly horizontal, and the yellow midrib more striking.
Grown as an indoor pot plant the trunks will be slender, the leaves longer and not so rigid, and the midribs will be light green. This is the form of the plant that I have grown indoors against a south-facing window for seven years. The plant has not been re-potted in that time, and is 75 cm tall. With the removal of dead leaves, the small tufts of green leaves on the slender trunks give the plant the appearance of a bonsai cabbage tree. I suggest its greatest use in future will be as an indoor pot plant competing for the place currently filled by costly stem propagated yuccas. Everyone could, indeed should have, a cabbage tree in their house!
Phenotypic variation of wild cabbage trees
 |
| Fig. 4 Cabbage trees (Cordyline australis) with flax (Phormium tenax) in a natural stand at Dipton, Southland. |
Their standardised form makes cabbage trees immediately recognisable, and yet each mature tree is strikingly different through infinite combinations of trunk dimensions, branching patterns, leaf size and shape, and arrangement of leaves in the tufts (Fig. 4). Clearly the environments where cabbage trees grow, and their age, strongly influence these variations. But is there a geographical pattern to this variation, and does the variation have a significant genetic component?
The late Dr Lucy Moore, arguably New Zealand's most notable woman botanist, studied and was fascinated by cabbage trees. She once said that if she was blindfolded and dropped off somewhere in New Zealand she would know what region of the country she was in if she saw only the form of the cabbage trees.
So with this observation in mind, measurements were made in the autumn of 1994 of trunk and leaf forms of 29 wild stands of cabbage trees at locations from Spirits Bay in the Far North to Dipton in Southland. These observations substantiated Lucy Moore's impressions (Harris et al. 1998). There were well-defined trends for stems becoming stouter the further south and the higher the altitude of the site of the stand, and leaf characters also showed changes along the latitudinal gradient.
Genotypic variation of cabbage tree seedlings
At the same time as the phenotypic characteristics of the cabbage tree stands were being recorded seed was collected from five trees of each stand. This seed was sown in August 1994 and the resulting seedlings were transferred to root trainers. In doing this we got the first clear indication that there were marked genetically based (genotypic) differences between cabbage tree populations and that these were related to geographical gradients.
One discovery was the occurrence of variation in red-brown pigmentation of the seedlings. This also showed a clear geographical pattern the incidence of red-brown colour increasing the further south the origin of the population. Related patterns were also shown for seedling height and leaf shape (Warwick Harris and Ross Beever unpublished data).
Genetic variation of cold tolerance
In spring 1995 the one-year-old seedlings of 28 of the cabbage tree populations were planted in the field at Mt Albert Auckland, Lincoln Canterbury (Fig. 5), and Invermay Otago. At each site the populations were each represented by 20 plants in a fully randomised layout. Since planting, measurements of the growth and other characteristics of the plants have been made at six-monthly intervals.
By a fortunate coincidence the 1996 winter at Invermay had an atypical cold spell with an extreme grass minimum of -9.7°C. This revealed clear differences in the freezing tolerance of the populations, those from the north showing severe damage and the most southern populations showing no damage. These differences were also shown at Lincoln, but not as markedly.
A consequence of the cold damage was the killing of the apical meristems of northern plants removing their dominance. Consequently at Invermay about half the plants, mostly from northern sites, have multiple stems, whereas plants of the southern populations are tall and single stemmed (Warwick Harris, Ross Beever and Bruce Smallfield, unpublished data).
 |
| Fig. 5 Five-year old cabbage trees (Cordyline australis) of different provenances in experimental garden cultivation at Lincoln in spring 1999. |
Flowering of cabbage trees
How long does it take for a cabbage tree to flower from the time it is sown? I have asked a good number of people this question and never received a definitive answer. From the cabbage tree population study it is clear that the answer is dependent on the conditions under which a plant is grown and its provenance.
The first of the trees flowered at Lincoln in spring 1998, four years after their seed was sown. Most interesting was that most of the plants that flowered were from the population from Dipton in inland Southland (Fig. 4). Fifteen of this population's 20 plants flowered whereas there was no flowering of plants from northern North Island locations (Warwick Harris, unpublished data).
Together with the cold tolerance response, this flowering response shows that genetically controlled physiological differences between cabbage trees are important in allowing C. australis to occur naturally over a wide geographical range.
This first flowering also revealed distinct differences in flower colour of cabbage trees varying from pure white and green to pink and brown pigmentation. As more plants flower it will be determined if the frequencies of these flower colours differ between populations.
Plant appearance
At Lincoln in spring 1999 some of the cabbage trees were close to being 4 m tall. Those that flowered in spring 1998 had branched from where the inflorescence emerged. But most were still single stemmed and these varied markedly in their height, the thickness of their trunks, the broadness of their leaves, the colour of their midribs, and the arrangement of their leaves (Fig. 5). Erect-leaved plants tend to have rolled leaves while floppiness is associated with flat leaves. As has been shown for harakeke-flax (King et al. 1996) rolling provides structural stiffness that allows the leaves to remain erect longer.
As the planting is fully randomised with each row of plants including 28 different C. australis provenances the similarity of form of plants within the provenances is not readily seen. Nevertheless I can walk through the plantation and reliably place plants as to their region of origin and often to their provenance.
Conclusions
The results from the cabbage tree provenance experiment have several applications:
- For their ornamental use, forms could be defined that would better suit plantings, for example as avenue trees, against tall or low buildings, as focal plants in landscape beds, or as potted patio plants.
- Where uniformity of plants is desired this could be achieved by using seed from defined provenances.
- More reliable extension of the use of cabbage trees as ornamental plants in colder climates could be obtained by the use of appropriate provenances. By this means the popularity of cabbage trees as a feature plant in Western Europe could be spread further.
- Should serious attempts be made to develop cabbage trees as novel sources of boutique fructose or vegetable oil, genetic variants exist that would enhance these uses.
- As cabbage trees are widely used in ecological restoration plantings, the results highlight the importance of using plants raised from local natural stands. This will reduce genetic contamination of local wild populations, and enhance the success of establishment, as the plants will be pre-adapted to the planting site.
- Although cabbage trees are still common and widespread, effort should be made to conserve representative wild populations across the species range. As indicated by C. 'Ti Tawhiti' and C. 'Karo Kiri', these populations may contain rare alleles in threat of extinction. More certainly these populations contain the genetic basis of the continuous variation that essential for the long-term vigour of cabbage trees. This vigour is needed for them to remain an icon of the New Zealand flora.
References
Harris, W. and Heenan, P. B. 1991: Cordyline 'Ti Tawhiti' and its relationship to Cordyline 'Thomas Kirk'. Horticulture in New Zealand 2(2): 2-5.
Harris, W. and Mann, J. D. 1994: Preliminary investigations of the suitability of Cordyline australis (Asphodelaceae) as a crop for fructose production. New Zealand Journal of Crop and Horticultural Science 22: 439-451.
Harris, W., Beever, R. E. and Heenan, P. B. 1998: Phenotypic variation of leaves and stems of wild stands of Cordyline australis (Lomandraceae). New Zealand Journal of Botany 36: 593-604.
Heenan, P. B. 1991a: Cordyline cultivar names — Three new combinations. Horticulture in New Zealand 2(1): 6-7.
Heenan, P. B. 1991b: A cultivar checklist for the New Zealand species of Cordyline (Asphodelacae). Horticulture in New Zealand 2(1): 8-12.
Heenan, P. B., Harris, W. and Bayliss, R. 1994: Cordyline australis 'Karo Kiri' (Asteliaceae): a new dwarf cabbage tree cultivar from New Zealand. Horticulture in New Zealand 5(2): 2-3.
King, M. J., Vincent, J. F. V., and Harris, W. 1996: Curling and folding of leaves of monocotyledons — a strategy for structural stiffness. New Zealand Journal of Botany 34: 411-416.
Potts, T. H. and Gray, W. 1870: On the cultivation of some species of native trees and shrubs. Transactions and Proceedings of the New Zealand Institute 3: 181-202.
Rutland, J. 1896: Traces of civilization: an enquiry into the history of the Pacific. Transactions and Proceedings of the New Zealand Institute: 29: 1-51.
Walsh, Rev. Canon 1900: On the occurrence of Cordyline terminalis in New Zealand. Transactions and Proceedings of the New Zealand Institute 33: 301-306.
![]()
- View
this paper as a PDF of the original proceedings

- Full
conference proceedings available as a hardcopy from the RNZIH
Reproduced from: New Zealand Plants and their Story
Proceedings of a conference held in Wellington, 1-3 October 1999
ISBN 0-9597756-3-3
Home | Journal
| Newsletter | Conferences
Awards | Join
RNZIH | RNZIH Directory | Links
© 20002026 Royal New Zealand Institute of Horticulture
Last updated: April 25, 2004